


Abstract
Objectives: Adhesion formation continues to be an important problem in gastrointestinal surgery. In recent years, methylene blue (MB) has been reported to be an effective agent for preventing peritoneal adhesions. However, its effects on the wound healing process are unknown. In the present study, we investigated the effects of MB on the early and late phases of anastomotic wound healing and on adhesion formation.
Methods: We randomly categorized 92 rats into 2 groups in bursting pressure measurements and 50 rats into 3 groups in the adhesion model. We divided the animals into saline-treated (n = 46) or MB-treated (n = 46) groups. Bursting pressures of the anastomoses were measured on postoperative days 3 and 7. In biochemical studies, tissue hydroxyproline levels, total nitrite/nitrate levels and nitric oxide synthase activity were measured on postoperative days 3 and 7. In the adhesion model, we randomly categorized rats into sham (n = 10), saline-treated (n = 20) and MB-treated (n = 20) groups, and the formation of intraperitoneal adhesions was scored on postoperative day 14. We compared the measurement of bursting pressure and biochemical measurements of tissue hydroxyproline levels, total nitrite/nitrate levels and nitric oxide synthase activity. Histopathological findings of specimens were presented.
Results: During the early phase of wound healing (postoperative day 3), bursting pressures, tissue hydroxyproline, total nitrite/nitrate levels and nitric oxide synthase activity in the MB-treated group were significantly lower than those of the saline-treated group. On postoperative day 7, there was no significant difference in these parameters between MB and saline-treated groups. In the adhesion model, MB caused a significant reduction in the formation of peritoneal adhesions.
Conclusion: MB prevents peritoneal adhesions but causes a significant impairment of anastomotic bursting pressure during the early phase of the wound healing process by its transient inhibitory effect on the nitric oxide pathway.
In abdominal surgery, the formation of intraperitoneal adhesions is a serious postoperative complication, validating relaparotomies in approximately 3% of all laparotomies performed. 1,2 Several earlier studies have developed different treatment strategies to reduce the incidence of intraperitoneal adhesions, including removal of fibrinous exudates by peritoneal lavage, use of proteolytic enzymes, anticoagulants, steroids, nonsteroid anti-inflammatory drugs, antihistamines, cytotoxic agents and use of substances such as olive oil or liquid paraffin.3–5 However, the benefits derived from such methods in relation to intraperitoneal adhesions remain unclear.
Since its discovery as the first synthetic antimalarial agent by Ehrlich in 1891, methylene blue (MB)6 has found uses in many different areas of clinical medicine, including treatment for septic shock, renal stones, methemoglobinaemia and encephalopathy. 7,8 In recent studies, it has been suggested that intraperitoneal application of MB can be used as an effective agent in the prevention of postoperative adhesions.9–11
Formation of adhesions occurs as a consequence of the normal physiological wound healing process.1–4 Following any trauma to the peritoneal surface, peritoneal mesothelial cells cover connective tissue containing blood vessels, collagen, lymphocytes, fibroblasts, macrophages, plasma cells and mast cells.3,4 Considering the physiological similarities between the healing process of intestinal anastomosis and the formation of adhesions, it is likely that agents affecting adhesion formation may also modulate the wound healing process. In the present study, we investigated the comparative effects of MB on anastomotic wound healing and adhesion formation models, and we discuss the mechanisms underlying the effects of MB on these processes.
Methods
The procedures followed in this study were in accordance with the Guide for the care and use of laboratory animals of the National Institutes of Health (Bethesda, Md.), and were approved by the Animal Ethics Committee of Ankara Oncology Training and Research Hospital. A total of 142 male Sprague–Dawley rats (270–320 g) were housed on a 12-hour light/dark cycle and at a temperature of 22°C–24°C. Before and after surgical procedures, the animals were provided with standard rat chow and water. One day before surgery, each rat was housed singly and was given water only 12 hours before the surgery. Sterile surgical protocols were maintained throughout the experiment. The rats were anaesthetized with IM ketamine (Ketalar, Parke Davis, Mich.; 50–40 mg/kg body weight) and xylazine (Rompun, Bayer AG, Leverkursen, Germany; 5 mg/kg body weight). Body weight, water and food intake were monitored daily throughout the postoperative period, for up to 7 days.
Anastomotic bursting pressure measurements
Following anesthesia, a 4-cm median laparotomy was made, and the left colon was transected (without resection of a segment) 4 cm proximally to the peritoneal reflection, taking care to preserve the marginal arteries. The bowel was restored by an end-to-end anastomosis with 6 interrupted, inverting sutures of 6/0 polypropylene (Prolene®; Ethicon Johnson & Johnson/ Ethicon Ltd., Istanbul, Turkey). Before closure of the abdominal wall, animals were assigned randomly to intraperitoneal application of 2 mL saline in the control group (n = 46) or 2 mL of 1% MB solution in the MB-treated group (n = 46). The abdominal wall was closed by continuous 3/0 polypropylene sutures.
On postoperative days (POD) 3 or 7 (n = 23 for each group), intra-abdominal adhesions were separated gently under a similar anesthesia. Healing of anastomotic wounds was evaluated by measuring anastomotic bursting pressure, which is reported to be preferable to other methods, such as bursting wall tension or tensile strength.12–14 POD 7 was the final time point for bursting pressure measurements, since beyond this, most colon segments rupture outside the anastomosis. 12,14 Briefly, 6 cm of colon segment centred by the anastomoses were resected and rats were sacrificed. Distal parts of the segments were closed with 2/0 silk sutures. The proximal parts of the segments were adapted to an intraluminal pressure manometer (monitoring kit L978-A07 Abott, Slingo, Ireland) and filled with isotonic NaCl solution with continuous infusion (4 mL/min). The bursting pressure (a peak pressure before anastomotic disruption) was measured with a pressure transducer (Peta, K 450, Ankara, Turkey). Since the evaluation of the bursting pressure might have caused damage along the anastomotic line, 3 animals from each group were spared for histopathological examination before the measurements. Tissue samples were kept in 10%-buffered formalin solution and stained with hematoxylin-eosin.
For biochemical measurements, the colonic anastomosis was cleared of overlying adherent tissue and excised with 0.5 cm of colon tissue on either side of the anastomosis. Cardiac blood was drawn in sterile fashion to determine serum total protein, albumin and NOS levels. Samples were either kept at −80°C for storage or were kept at −25°C and evaluated immediately for biochemical parameters.
Measurements of nitrite and nitrate levels
Total nitrite and nitrate content (NOx), both stable end products of nitric oxide, were measured spectrophotometrically with a nitric oxide assay kit (Calbiochem, San Diego, Calif.). Tissue samples were pooled, weighed and homogenized in phosphate buffered saline (PBS) at pH 7.4 (10 vol) with a polytron (PT 1020, 350D, Lucerne, Switzerland). The homogenate was centrifuged at 10 000 × g for 20 minutes, and supernatant was filtered through 10 k of MW cut off Ultrafree-MC (Millipore, Bedford, Mass.). Filtrates were incubated with nitrate reductase for 3 hours and assayed for nitrite contents with Griess reagents and measuring absorbance at 540 nm.
Measurements of nitric oxide synthase activity
Nitric oxide synthase (NOS) activity was determined by measuring the conversion of [3H]l-arginine to [3H]L-citrulline with an NOS assay kit (Calbiochem, San Diego, Calif.). Anastomotic tissue samples were homogenized in 20 volumes of 25 mM Tris-HCl, 1 mM EDTA, 1 mM EGTA, pH 7.4. The homogenates were centrifuged at 10 000 × g for 5 minutes, and at 4°C supernatant was collected. Briefly, 5 μl of supernatant for each sample was incubated with a buffer containing cofactors required for NOS enzyme activity (25 mM Tris-HCl, 1 mM NADPH, 3 μM tetrahydrobiopterin, 1 μM flavin adenine dinucleotide, 1 μM flavin adenine mononucleotide, 0.8 mM CaCl2, and 0.05 μC [3H]l-arginine, specific activity, 60 Ci/mmol) for 60 minutes at 35°C. Reaction was stopped with a stop buffer (400 μl) composed of 25 mM Tris-HCl, 2.5 mM EDTA, pH 5.5, and 100 μl equilibrated resin was added. Samples were transferred to spin cups and centrifuged at 1200 g for 30 seconds. The radioactivity in eluates was quantified in a liquid scintillation counter. NOS activity was determined from the difference between [3H]-L-citrulline produced from control samples and containing 1 mM L-NAME, a competitive inhibitor of NOS. Protein content in homogenates was determined with a protein assay kit based on the Lowry method (Bio-Rad, Hercules, Calif.), and NOS activity was expressed as pmol of [3H]-L-citrulline formed per min/mg protein.
Measurements of hydroxyproline levels
Hydroxyproline levels are shown to indicate the amount of collagen in tissues, 15 and a direct relation between anastomotic healing and tissue hydroxyproline levels has been reported. 15–17 The tissues (30–50 mg) were placed into hydrolysis tubes. Fifty mM potassium phosphate buffer pH 7.0 and an equal volume of concentrated HCl were added to each tube, and the samples were hydrolyzed at 110°C for 16 hours. The samples were oxidized with Chloramine-T solution (pH 8.5), and the Ehrlich’s reagent was added. The colour was allowed to develop at 60°C for 25 minutes, and the absorbency at 560 μm was determined with Bergman and Loxley’s method.18 Total protein on tissue homogenates was determined by the addition of trichloroacetic acid (TCA, 10% final concentration) to precipitate proteins, and the sample was centrifuged at 2500 g for 10 minutes. The amount of protein in sediment was determined with a protein assay kit based on the Lowry method (Bio-Rad, Hercules, Calif.).
Adhesion studies
Following anesthesia, median laparotomy was made 4 cm in length, the small bowel was retracted upward and the cecum was exposed. The adhesions were induced by scraping the wall of the cecum to cause punctuated serosal hemorrhages. Rats were randomly assigned to sham-operated (n = 10), saline-treated (n = 20) and MB-treated (n = 20) groups. Shamoperated rats had a laparotomy but no induction of adhesions. After laparotomy in saline-treated and MB-treated groups, 2 mL of saline and 2 mL of 1% MB solution were instilled into the peritoneal cavity, respectively. The abdominal cavity was closed with continuous 3/0 polypropylene sutures. After 14 days, relaparotomies were performed, and the abdominal cavity was inspected through a U-shaped incision. The adhesions were identified, counted and graded by 2 independent investigators who were blinded to the assigned groups with the classification described by Nair and colleagues19 (Table 1).
Adhesion score categories according to Nair and others19
Data analysis
The variances of the bursting pressures and hydroxyproline levels, but not the adhesion index, were homogeneous, and the distributions were normal. Therefore, we used the Student’s t test in the wound-healing model and the Kruskal–Wallis and Mann–Whitney U tests in the adhesion model to evaluate the differences among the groups. The criterion for significance was accepted as p < 0.05.
Results
Effects of MB on anastomotic wound healing
During the course of experimental protocols, no animal in any of the treatment groups died. There were no wound infections as assessed by clinical inspection. On POD 1–3, animals from all groups demonstrated weight loss (Fig. 1). However, there were no significant differences between the groups at any time point (n = 12 for each group; p > 0.05). The means of the metabolic and nutritional parameters measured in saline-treated and MB-treated animals within 3 postoperative days are presented in Table 2. There were no statistically significant differences between the means of these parameters measured in saline and MB-treated groups, respectively (n = 12 for each group; p > 0.05).

Postoperative weight changes in animals treated with either saline or methylene blue (MB). Both saline- and MB-treated animals experienced similar weight loss during the postoperative healing period. There was no statistically significant difference between the groups at any time point (n = 12, p > 0.05).
Some of the metabolic and nutritional parameters in saline and MB-treated groups measured during postoperative 3 days
Compared with the saline-treated group (controls), MB treatment resulted in statistically significant reduction in the means of anastomotic bursting pressure measured on POD 3 (51.5, standard error of the mean [SEM] 4.7 mm Hg v. 70.4, SEM 3.9 mm Hg for controls; p = 0.001; Fig. 2A). Interestingly, MB treatment did not cause a significant alteration on the means of the bursting pressures measured on POD 7, when compared with controls (167.2, SEM 4.7 mm Hg v. 170.8, SEM 5.1 mm Hg for controls; p > 0.05; Fig. 2B). Additionally, we measured the circumferences of the anastomotic lines after bursting of the anastomoses. Compared with controls, the differences in the length of circumferences between the groups were not significant either on POD 3 or 7 (data not shown), suggesting that tissue edema was not a complicating factor in the bursting pressure measurements.

Effects of methylene blue (MB) treatment on anastomotic bursting pressure measured on postoperative days (POD) 3 and 7. Animals received intraperitoneal application of either 2 mL saline (n = 23) or 2 mL MB (n = 23). Measurements of anastomotic bursting pressures on POD 3 and 7 are presented in A and B. *p = 0.001.
Hydroxyproline levels of samples taken from the anastomotic line on POD 3 indicated that there is a statistically significant difference between the control and the MB-treated group (mean 3.1, SEM 1.2 mg/g tissue v. mean 3.7, SEM 1.4 mg/g tissue for controls; p = 0.016), which suggests an impaired collagen synthesis in this group of animals. Similar to earlier findings on bursting pressures, there were no statistically significant alterations on the hydroxyproline levels measured on POD 7 (mean 5.4, SEM 0.2 mg/g tissue in the MB-treated group v. mean 5.3, SEM 0.3 mg/g tissue for controls; p > 0.05). Total protein contents in homogenates of anastomotic tissue samples were not significantly different between the groups either on POD 3 (mean 14.3, SEM 2.6 mg/g tissue v. mean 13.8, SEM 2.4 mg/g tissue for controls; p > 0.05) or POD 7 (mean 17.4, SEM 3.1 mg/g tissue v. mean 16.7, SEM 2.9 mg/g tissue for controls; p > 0.05). This suggests that the alterations in bursting-pressure and hydroxyproline levels measured at POD 3 cannot be ascribed to total protein synthesis and/or weight loss.
Effects of MB on NO pathway
Because MB is a known inhibitor of nitric oxide synthase,20 we investigated the effect of MB treatment on total nitrite and nitrate levels (NOx) on anastomotic tissue and plasma samples. Compared with samples from the saline-treated group, MB treatment caused a significant inhibition of NO production as measured by NOx levels in anastomotic tissue samples on POD 3 (mean 34, SEM 4 μM v. mean 62, SEM 7 μM for controls; n = 12, p < 0.05). However, tissue NOx concentrations measured on POD 7 were not altered by MB-treatment (mean 35, SEM 4 μM v. mean 38, SEM 5 μM for controls; n = 12, p > 0.05). Plasma NOx levels remained unchanged throughout the postoperative healing period. Results of NOx measurements of tissue and plasma samples in control and in MB-treated groups for POD 3 and 7 are presented in Figure 3.
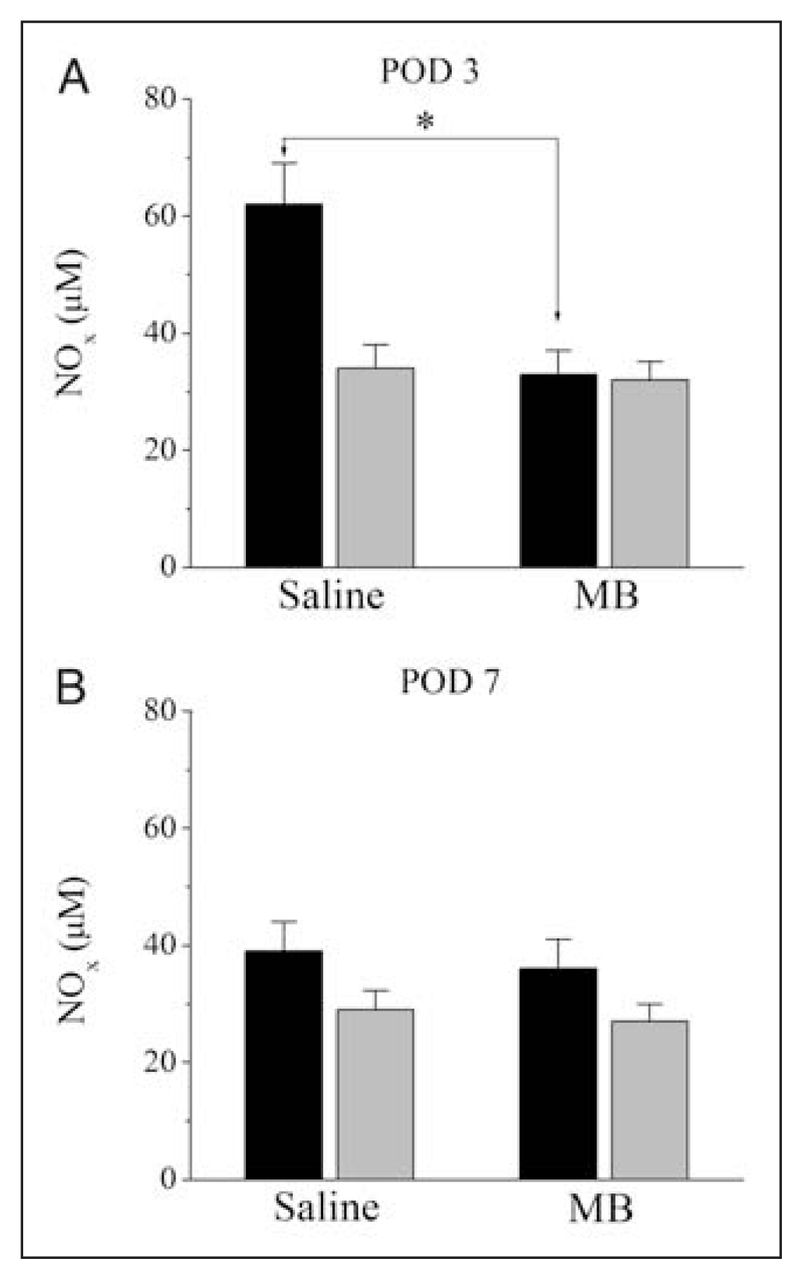
Effects of methylene blue (MB) treatment on total nitrite and nitrate levels measured in anastomotic tissue and plasma samples on postoperative days (POD) 3 and 7. Total nitrite and nitrate levels were measured with Griess reagent. Animals received intraperitoneal application of either 2 mL saline (n = 12) or 2 mL MB (n = 12). Measurements of total nitrite and nitrate levels on POD 3 are presented in panel A and on POD 7 are presented in panel B, respectively. *p = 0.001. The black and grey bars indicate tissue and plasma levels. NOx = total nitrite and nitrate content.
Effects of MB treatment on NOx concentrations observed on POD 3 may be due to MB inhibition of NOS activity. Thus, we have measured NOS activity on POD 3 from anastomotic tissue samples. Compared with the saline-treated group, NOS activity decreased significantly in samples from the MB-treated group (mean 5.3, SEM 2.1 v. mean 12.4, SEM 3.6 pmol [3H]-citrulline/ mg protein/min for controls; n = 12, p < 0.05). The results of NOS activity measurements on tissue samples in the control and MB-treated groups are presented in Figure 4.
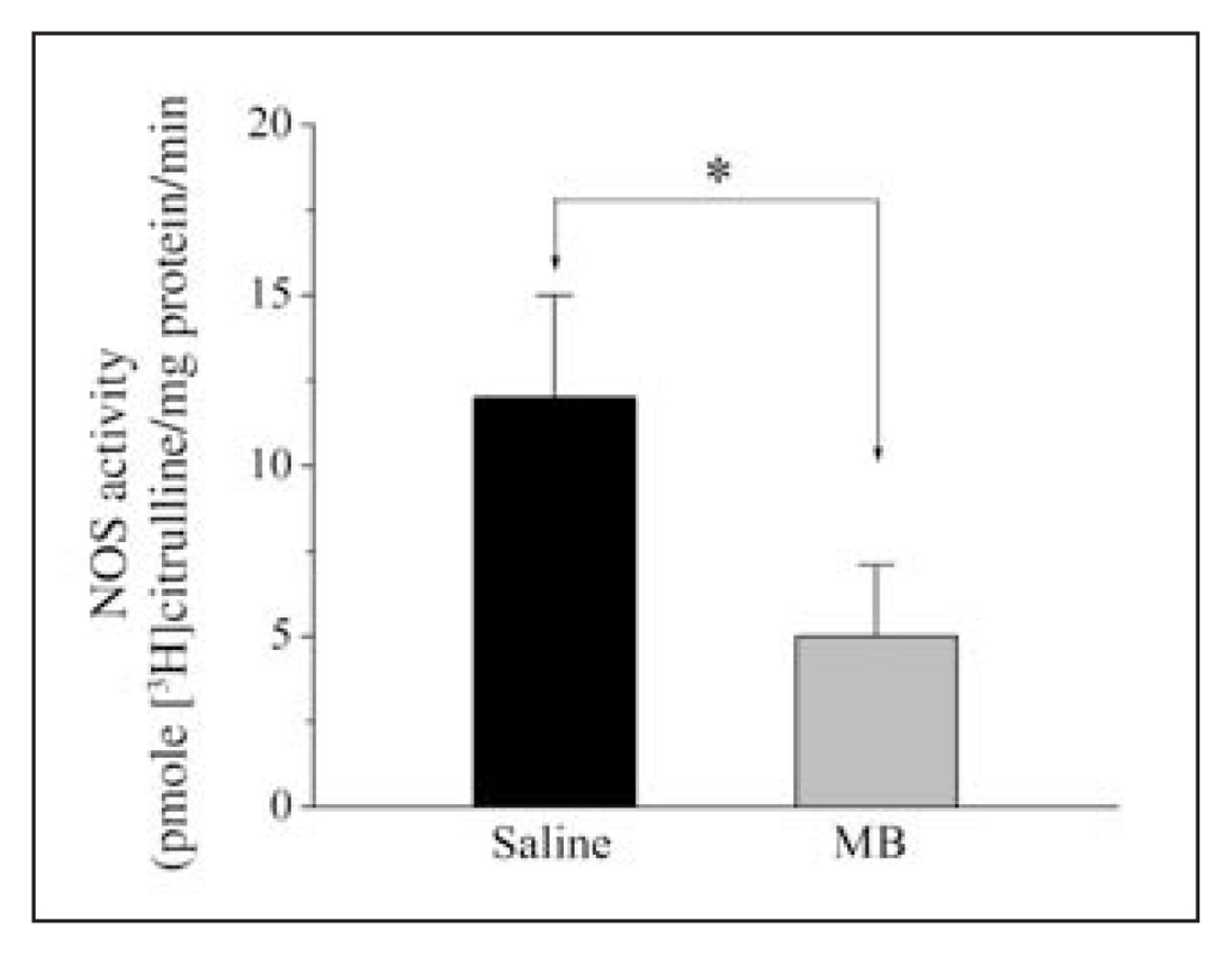
Effects of methylene blue (MB) treatment on nitric oxide synthase (NOS) activity measured in the rat colonic anastomoses on postoperative days (POD) 3 and 7. NOS activity was measured by determining the rate of conversion of labelled arginine to citrulline in anastomotic tissue samples Animals received intraperitoneal application of either 2 mL saline (n = 12) or 2 mL (MB) (n = 12). Measurements of NOS activity on POD 3 and 7 are presented in panels A and B. *p = 0.0001.
Effects of MB on adhesion formation
Results of experiments summarizing the semiquantitative grading of adhesion severity in the adhesion formation model are presented in Figure 5. The mean adhesion score of animals in the sham-operated group was 0.5, SEM 0.5 (n = 10). The adhesion score of animals in which adhesion had been induced and treated with saline was mean 3.8, SEM 0.4 (n = 20). The adhesion score of animals in adhesion-induced and MB-treated groups was mean 1.1, SEM 0.3 (n = 20). The difference of the mean values between saline and MB-treated groups was statistically significant (p < 0.0001).

Effects of methylene blue (MB) treatment on experimentally-induced adhesion. Animals received either no intraperitoneal application (sham; n = 10), 2 mL saline (control; n = 20) or 2 mL MB (MB; n = 20). Mean values of adhesion scores (± standard error of the mean) are presented. *p = 0.001.
Findings of the histopathological examinations are presented in Figure 6. Figure 6A is the anastomotic control group. In Figure 6B, there is poor anastomotic wound healing in the MB-treated group. There is a significant gap on the anastomotic line, and the fibroblastic activity is minimal. The submucosal layer under the anastomotic line is thin, and the omentum covers the anastomotic line, probably due to minimal leakage from the anastomosis.
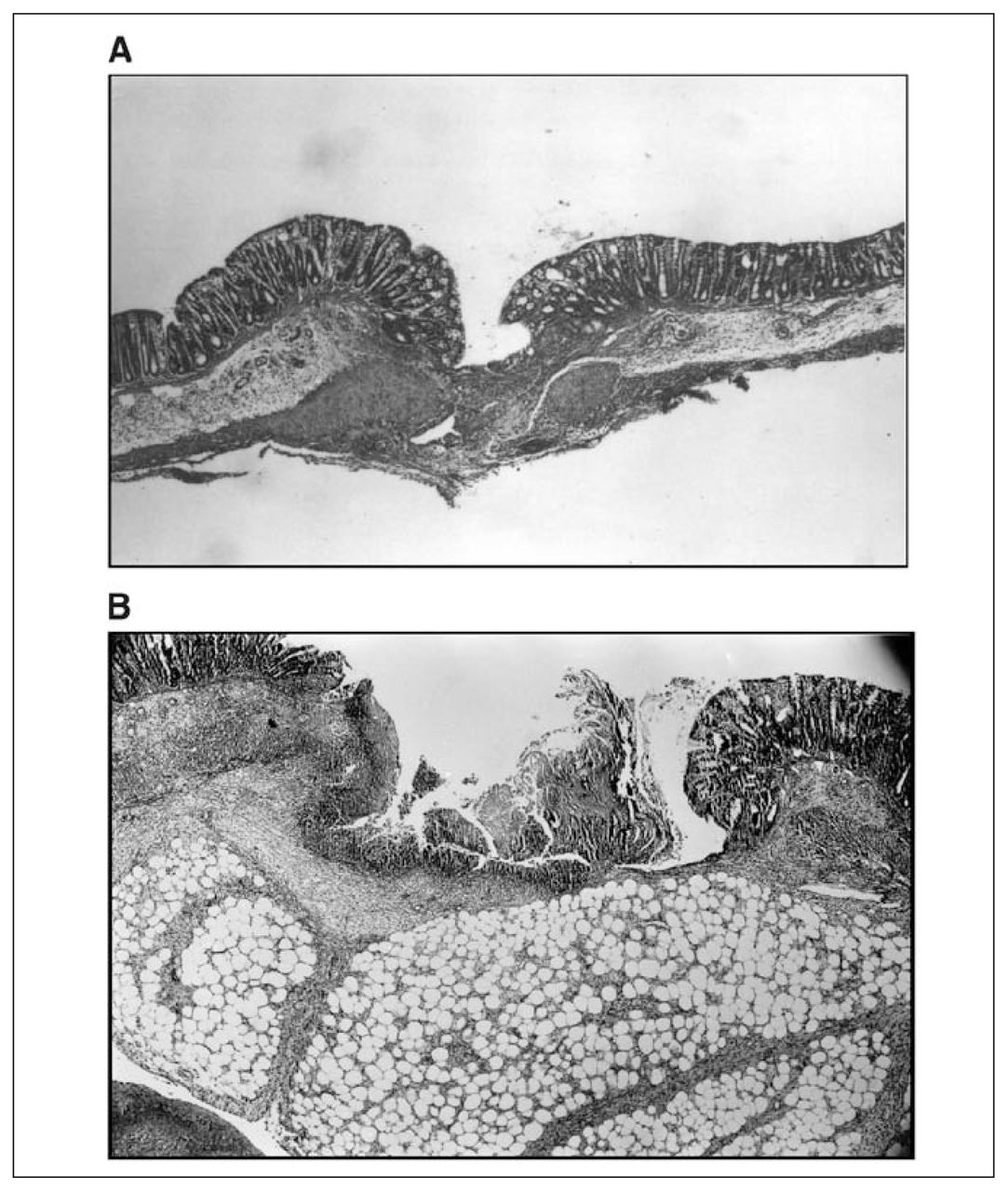
Histopathological examinations of the effect of methylene blue (MB) treatment on adhesion and anastomotic wound samples. A) Hematoxylin-eosin stained photomicrograph (20× magnification) from anastomotic control group. The anastomotic line is at the centre of the figure. The fibroblastic activity has begun on the subseroza and muscularis propria. The continuity of the intestine is not damaged. There is no gap between subseroza and muscularis mucosa. B) Hematoxylin-eosin stained representative photomicrograph of anastomotic wound in the MB-treated rats on the third postoperative day (20× magnification). Impaired anastomotic wound healing with a significant gap and necrotic area is noticeable.
Discussion
Results of the present study provide the first indication that, although intraperitoneal application of MB effectively reduces the formation of adhesions, it also causes significant impairment of anastomotic wound healing during the early phases of the wound healing process.
Earlier studies with intraperitoneal MB applications reported that optimal effective MB concentrations are in the range of 1%–2% (v/v).9,10 Consistent with these results, intraperitoneal application of 1% MB reduced the formations of adhesions significantly without causing toxicity in the animals. However, MB was ineffective in preventing adhesion formations at higher concentrations (5%–7%), and no toxicity was evident.10,21,22 Similarly, low concentrations of MB (0.1%–0.5%) were ineffective in decreasing adhesion formations. 10,23 Although there was acute weight loss following colon anastomosis, there was no significant difference in weight loss between the control and the MB-treated groups. Further, there were no significant alterations on the metabolic and nutritional parameters measured in our study between saline-treated and MB-treated animals (Table 2). Therefore, the difference observed in anastomotic healing, tissue hydroxyproline and NOx levels on POD 3 could not be ascribed to metabolic conditions, weight loss or toxic effects of MB treatment.
The mechanism of actions of MB on peritoneal adhesion is currently unknown. MB has been shown to be a nonselective NO synthase inhibitor; that is, it acts on both constitutive NO synthase isoforms as well as on inducible NO synthase isoforms.20 Because of this action, it is likely that the effect of MB on adhesion formation is mediated by the inhibition of the NO pathway. However, NO synthase inhibitors such as N(G)-nitro L-arginine methyl ester (L-NAME) do not mimic the effects of MB on surgically-induced adhesion models. 9,24 Paradoxically, NO donors such as L-arginine and spermine-nitric oxide (SPERNO) decrease the formation of adhesions.22,24,25 Thus, the pharmacological profile of the effects of MB on adhesion formation is not consistent with the inhibition of the NO pathway. MB has several other biological effects that could mediate its inhibitory actions on adhesion formation. For example, as an electron acceptor, MB inhibits the generation of oxygen radicals, such as superoxide, by competing with the molecular oxygen for the transfer of electrons from xantine oxidase.26,27 In fact, several compounds with antioxidant properties including vitamin E, super oxide dismutase, catalase and allopurinol, have been effective in preventing adhesions in earlier studies.28,29
Despite the salient effects of MB on adhesion formation, the early phases of surgically-induced wound healing were impaired significantly by intraperitoneal application of MB. To our knowledge, this is the first report on the effect of MB on the anastomotic wound healing process. The mechanisms of action of MB on wound healing are currently not well defined. However, several lines of evidence suggest that inhibition of the NO pathway plays a role on the observed effects of MB during the early phases of wound healing.30 NO is synthesized in wounded tissue, and the temporal pattern of its synthesis correlates with the wound healing process.31,32 Both wound-derived macrophages and fibroblasts express the inducible form of NO-synthase (iNOS).30 It has also been reported that the synthesis of NO reaches maximal levels on the third POD.30–33 This period corresponds to the inflammatory phase of the wound healing process. Interestingly, most of the primary effects of NO, including vasodilatation, antimicrobial activity, antiplatelet aggregation activity and induction of vascular permeability, are especially relevant to the inflammatory phase of the wound healing.30–32 As well, NO stimulates wound healing by increasing collagen formation in dermal fibroblasts. 30 Inhibition of iNOS by competitive inhibitors decreases collagen deposition and breaking strength of incisional wounds.30 NO has also been shown to increase angiogenesis, and eNOS inhibitors impair angiogenesis in granulation tissue.30,34 NOS inhibitors cause significant impairment on anastomotic wound healing,31,35,36 and these agents are maximally effective during the inflammatory phase of wound healing. 30,32,34 It is likely that, during the early phase of wound healing (POD 3 in this study), the effects of MB on anastomotic bursting pressure is mediated by the inhibition of the NO pathway. In line with this hypothesis, our results indicate that both NOx content and NOS activity in the anastomotic tissue samples of MB-treated animals were decreased significantly on POD 3. On the basis of our results and on earlier literature, it is likely that MB inhibits the early phases of wound healing by inhibiting NOS activity and decreasing NOx levels in anastomotic tissue.
Although MB impaired the early phases of wound healing, it did not cause significant alterations on anastomotic bursting pressure, tissue hydroxyproline levels, tissue NOx content and NOS activity measured on the seventh POD. In earlier studies, it was found that treatments of animals with colonic anastomoses by NOS inhibitors including L-NAME and lysine (L-NIL) activated adaptive mechanisms that mainly increased expression of cytokines and growth factors.35 In a recent study in NOS knockout mice,37 the number of iNOS-expressing cells increased significantly on POD 7 but remained unaltered on POD 3 in the incisional wound model. Such transient nature of the MB effect on NOx levels and anastomotic wound healing may be due to compensatory changes on tissue NOx levels by POD 7.
In contrast to tissue levels, plasma concentrations of NOx have been found to remain steady throughout the post-wounding period.33 MB is a cationic dye with a pKa value of 0–1 and is ionized completely in physiological solutions.38 Thus, it is not absorbed easily and could remain in injected areas for a week.39 It is possible that MB lacks any effect on serum NOx levels on both POD 3 and 7, owing to its decreased bioavailability in circulation.
Clinically, the application of MB during surgery would be appropriate, because the adhesion formation is triggered at early stages.2,4,5 However, due to adverse effects of MB on anastomotic wound healing, the use of MB in anastomotic procedures would not be suitable. Conversely, MB can be used as an adjuvant in abdominal surgeries involving extraintestinal interventions, including postoperative adhesions, extra-intestinal tumours, hernias and intra-abdominal abscess. Among these conditions, adhesions present a widespread problem and develop after nearly all abdominal surgical procedures (i.e., open or laparoscopic).40,41 Intestinal obstruction is the most severe consequence of adhesions. Of patients who require abdominal reoperation, 30%–41% have adhesion-related intestinal obstruction. For small-bowel obstruction, the proportion rises to 65%–75%.40,41 In a recent clinical study, a bioresorbable adhesion barrier has also been suggested to prevent adhesion formations with increased risk of sequelae associated with anastomotic leak.42 We believe that further clinical research with MB, a commonly available and relatively nontoxic compound, is needed to investigate the effectiveness of MB on a selected patient population.
Results indicate that, although MB decreases the formation of intraperitoneal adhesions, it causes a significant impairment on anastomotic bursting pressure and collagen synthesis during the early phases of wound healing through its effects on the NO pathway. Considering these deleterious effects of MB on the wound healing process, we suggest that the use of this agent in the prevention of adhesion formation requires careful clinical monitoring during the early phases of wound healing.
Acknowledgements
We thank Ms. Mary Pfeiffer of NIDA/NIH for her helpful reading of the manuscript.
Footnotes
This experimental study was carried out in the Laboratories of the Animal Facilities and Department of Pathology, Ankara Oncology Training and Research Hospital, and Department of Biochemistry, University of Hacettepe, Ankara, Turkey.
Competing interests: None declared.
- Accepted January 28, 2005.
References
In this issue
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Article tools





Related Articles
Cited By...
- No citing articles found.